Hay muchos tipos de membranas celulares. Son algo más que
sistemas de la célula:
- Forman o aíslan compartimentos cerrados.
- Regulan el paso o movimientos de sustancias a través de ellas.
- Se encargan del paso de información: las membranas tienen receptores específicos de un ligando.
- Se encargan del reconocimiento intercelular.
- Son la plataforma donde se van a ordenar los componentes de un determinado proceso.
- Se encargan de transducciones de energía y transformaciones de energía.
- Participan en la transmisión del impulso nervioso.
- Se encargan de la absorción de sustancias.
- Etcétera.
La membrana es una estructura estudiada a finales del siglo
XIX. Se basaron en observaciones en las que se veía que los compuestos
liposolubles atravesaban mejor la membrana que las sustancias hidrosolubles.
 |
| Con la microscopía electrónica se consiguió una visualización de la membrana, aunque algo artefactual. Los primeros modelos eran algo erróneos. Lysec estudió la membrana usando un glóbulo rojo. Esta célula solo presenta membrana plasmática, careciendo de núcleo y orgánulos. Por eso se sigue usando mucho para el estudio de membranas. Se observó que, si se disolvía la membrana y se ponía en medio acuoso, aparecía una capa de naturaleza grasa. Prácticamente, la membrana disuelta ocupaba el doble, aproximadamente, de la superficie total del eritrocito. La membrana debía ser, por lo tanto, doble. Se trata, por lo tanto, de una bicapa. |
Con microscopía electrónica se observa una estructura
similar a la siguiente:

Se postuló que la membrana no era solamente una bicapa
lipídico. Se vio rápidamente que aparecían proteínas asociadas. Estas darían
cohesión a la membrana y facilitarían el paso de compuestos hidrófilos.
Danielli propuso una membrana que estaría constituida por
una bicapa lipídica envuelta o tapizada por una envuelta protéica, de forma que
las proteínas constituirían las zonas electrondensas.
 |
| Membrana de Danielli |


 |
| Modelo del mosaico fluido |
Las proteínas periféricas son hidrófilas. Se unen a la
membrana por interacciones hidrofílicas con las cabezas de los lípidos o a las
partes hidrófilas de las proteínas integrales.
Las proteínas integrales están unidas a la bicapa lipídica.
Son en gran parte hidrófobas. Están sujetas por interacción hidrófoba.
Dentro de las proteínas integrales las hay transmembrana, si
la atraviesan de parte a parte. Es frecuente que se formen en las integrales,
poros hidrófobos.
No hay enlaces covalentes. Se basa en la cooperación de
muchos enlaces débiles.
Los hidratos de carbono siempre están hacia el exterior,
nunca hacia el citoplasma.
Se mantiene la existencia de una bicapa lipídica y de un
contenido protéico. En el caso de las proteínas, se cambia bastante su
posición. Muchas proteínas están sumergidas, total o parcialmente, en la
bicapa. Se sabe que existen proteínas que se encuentran por la parte exterior
de la bicapa, enlazadas con los lípidos de la membrana o con otras proteínas.
Además, en la cara exterior o
extracitoplasmática, la que mira al exterior de la célula, existe un
componente glucídico. Este modelo explica mejor las características físico –
químicas de la membrana.
Hay una gran
movilidad en la bicapa, no solo de las proteínas, también de los lípidos. No
son estáticos.
Falta otro detalle. Las fibrillas que se ven en microscopía
electrónica. Se trata del componente glicídico, los polisacáridos unidos sobre
todo a ciertas proteínas, formando un entramado. Algunos lípidos presentan
también componentes glucídicos.
Las membranas, por lo tanto, son asimétricas. Y una de estas
asimetrías es la disposición de los árboles de azúcares, que como acabamos de
decir están solo en una de las caras de la membrana. En el caso de la membrana
plasmática, ya indicamos que están hacia el exterior. Cuando están muy
desarrollados, los árboles son muy grandes, hablamos de glucocálix. En las
membranas internas, como el aparato de Golgi, o en vesículas interiores, los
azúcares se sitúan hacia la cara interior o luminar de las mismas.
En las células, en el interior de cloroplastos o
mitocondrias, no hay hidratos de carbono. En las bacterias, a veces, los
lípidos de una monocapa no corresponden con el de la otra monocapa.
Los fosfolípidos tampoco son simétricos. En la membrana de
los eritrocitos, en la cara exterior, encontramos fundamentalmente
fosfatidilcolina. En la interior hay otros fosfolípidos.
Las proteínas transmembrana tienen una orientación fija,
siempre es la misma. En el eritrocito, la glicoforina siempre se coloca de la
misma manera.
Las proteínas de membrana se sintetizan en los ribosomas que
están pegados a la membrana. La conformación tiene que ver con el transporte a
través de la membrana.
Un nuevo dato microscópico vino a solucionar problemas y a
confirmar el nuevo modelo. Cuando preparamos imágenes por criofractura, las
muestras se parecen más a la realidad, ya que no se llevan a cabo inclusiones
ni fijaciones. Con este tipo de microscopía se ve una membrana distinta. Hay
dos capas similares, que corresponden a las dos hemicapas lipídicas, con una
zona intermedia más frágil. En muchas ocasiones la célula, en lugar de partirse
por la mitad, se separan las dos hemimembranas. Se compraba que son muy
parecidas, que presentan una alternancia de zonas lisas y zonas con huecos o
estructuras que sobresalen. Además, estos son complementarios. La explicación
que se da está en relación con el mosaico fluido, las dos hemimembranas
corresponden a dos partes de la membrana lipídica. La zona que separa las dos
hemimembranas son las zonas con menos enlaces, son poco resistentes, lo que
facilita que en la criofractura sea por esa zona por donde se rompa, por donde
se separe. Por otro lado, cuando se separa, cada lado tira de sus proteínas.
Las proteínas exteriores no tienen problema, pero las que están dentro de la
bicapa se van hacia uno de los dos lados, quedando un hueco en el lado del que
son arrancadas y un saliente en la zona donde se queda enganchada la proteína.
 |
| Imágenes de criofractura |
 |
| Vaina de mielina de los axones |
Se ha llegado a la conclusión de que, debido a la fijación,
exclusión y resto de técnicas agresivas previas a la microscopía electrónica,
se produce una desorganización de la membrana plasmática, con pérdida del
componente protéico intermembrana. De ahí la imagen errónea del microscopio
electrónico de transmisión (sin técnica de criofractura).
La membrana tiene, por lo tanto, un componente lipídico, un
componente protéico y un componente glucídico.
Todas las membranas estudiadas tienen una estructura
semejante, con una composición de alrededor de un 40% de lípidos y un 60% de
proteínas, pudiendo llegar a estar incluso invertidos estos datos, es decir,
60% de lípidos y 40% de proteínas.
Fosfolípidos.
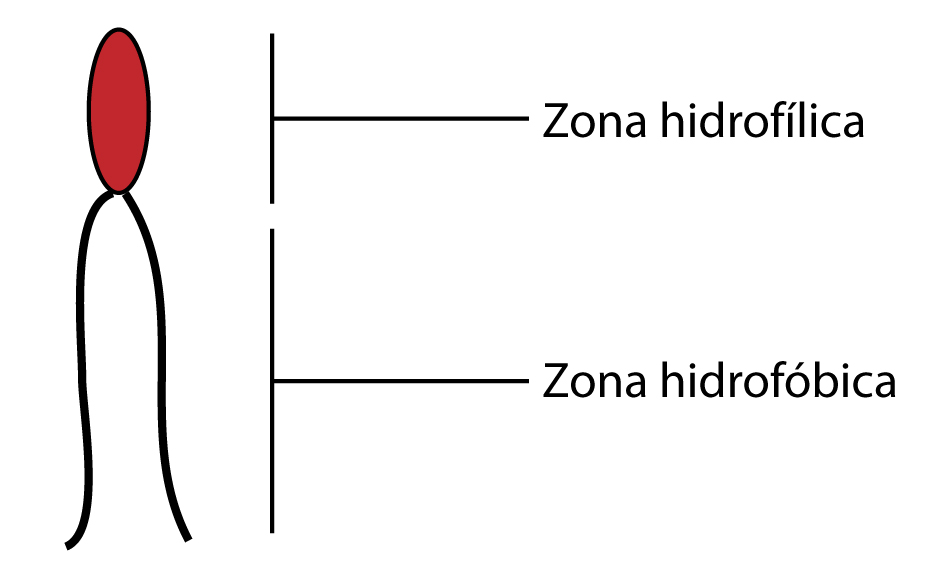 |
| Fosfolípido |
Los lípidos de la membrana biológico están en un grupo muy
característico, pertenece a los fosfolípidos. En el caso de eucariotas, está
también el colesterol.
Los lípidos tienden a esconder la cola hidrfóbica, formar
micelas o formar bicapas protectoras.
 |
| Esquema del colesterol |
Para los fosfolípidos y glucolípidos, la estructura más
estable es la bicapa. Se adaptan espontáneamente. Las bicapas forman vesículas.
Se les llama liposomas. Las fuerzas que las estabilizan son las interacciones
hidrófobas entre las colas y las interacciones de Van der Wallas al estar muy
cerca. Por otro lado, las cabezas polares pueden formar puentes de hidrógeno o
puentes salinos con el medio acuoso.
 |
| Fosfolípido |
Muchas membranas tienen muchos tipos de fosfolípidos. Por
ejemplo, en algunas aparecen los esfingolípidos, que son lípidos que derivan de
la esfingosina. En los esfingolípidos el grupo polar estará constituido por
algún hidrato de carbono. Es decir, las dos moléculas hidrófobas provienen, una
de la esfingosina y otra del ácido graso.
 |
| Esfingosina |

Esta estructura en lámina debe ponerse en un medio que aísle el agua. por ejemplo, en forma de anillo. Lo que ocurre es que se cierra la bicapa en una forma esférica. Esta forma de cerrarse es lo que se supone que sucedió en las primeras células. Se creó una estructura en la que hay un medio interno. Y sin gasto de energía, se consigue tener una superficie que, espontáneamente, aísla el interior del exterior.
Además, la membrana es un medio muy aislante, solo es atravesada por sustancias liposolubles. Por otro lado, se puede variar el aislamiento, por medio de compuestos o estructuras que la atraviesen. Se puede perforar se cerrará automáticamente, de forma espontánea, sin gasto de energía. El paso de sustancias se lleva a cabo gracias a proteínas. El sistema de ruptura y reposición de la membrana es aprovechado por el proceso de exocitosis y endocitosis, que sirve para introducir o sacar grandes cantidades de sustancias.
Los lípidos son de composición variable. Los lípidos de la
membrana son además moléculas con una movilidad elevada. Se han demostrado
estos movimientos de los fosfolípidos mediante resonancia de espín electrónico.
Se marcan determinados lípidos con grupos químicos con un electrón desapareado.
Y así vemos que los lípidos tienen movimientos de difusión lateral, de rotación
sobre su eje y movimientos de inversión en la membrana o movimiento flip-flop.
Este último movimiento es casi inexistente, con excepción de ciertas zonas de
membrana como la membrana del retículo endoplásmico, debido al mecanismo de
fabricación de la membrana. Los movimientos laterales, en cambio, se realizan
sin problemas. Cada célula modifica su membrana para que los movimientos no se
hagan de forma azarosa, sino ordenadamente.
 |
| Movimientos de los fosfolpidos |

Otra propiedad importante de las membranas será su fluidez.
Por una parte está la fluidez de los propios lípidos. Tienen varios tipos de
movimientos dentro de la bicapa.
Los lípidos se pueden desplazar rápidamente dentro de una
misma monocapa y conservando su orientación.
Hay factores
que influyen en la fluidez de la membrana. A más temperatura, más fluidez y
viceversa. Se habla de temperatura de transición, que sería la temperatura en
la que la membrana pasa de tener una consistencia líquida a pasa a una
consistencia de gel cristalino. Cada membrana tendrá su propia temperatura de
transición.
Otros factores serán los factores de composición. Son
fundamentalmente tres:
- Grado de saturación de los ácidos grasos. Cuando hay un mayor número de ácidos grasos con dobles enlaces, la cadena hace quiebros, las colas tienen dobleces en lugar de ir rectas, provocándose huecos. Esto hace que la membrana sea más fluida y su temperatura de transición más baja.
- Ácidos grasos de cadena corta. Se dificulta el empaquetamiento por lo que la membrana se hace más fluida. Al igual que en el caso anterior, se crean huecos que provocan que las fuerzas de Van der Waals sean menos efectivas.
- Presencia de colesterol: solo aparece en eucariotas, nunca en procariotas. Modula la fluidez. A temperatura alta, el colesterol evita el exceso de fluidez. Y viceversa, cuando la temperatura es baja, evita la cristalización. Hace un efecto de unión, acerca las moléculas. Por otro lado, separa las moléculas disminuyendo la fluidez. Es decir, depende de como se coloque. Hace que los cambios de temperatura no se hagan sentir en exceso.
Si las
membranas están compuestas por un tipo de lípidos, cuando baja la temperatura,
llegaremos a conseguir una rigidez en la bicapa. Si hay varios tipos de
lípidos, se formarán islas, es decir, los más fluidos se van uniendo entre si.
En las membranas biológicas esto no ocurre, porque suelen poseer una cola
saturada y una cola insaturada, lo que hace que las características se repartan
y no aparezcan islotes. Debemos tener en cuenta que estos islotes provocarían
la ruptura de la membrana. De esta forma, las células animales pueden aguantar
temperaturas de entre -20ºC hasta +50ºC.
La célula puede solventar los problemas de fluidez de muchas
maneras. Los organismos inferiores cambian la composición de las colas de la
membrana. En eucariotas no hay un control de este tipo. Pero encontramos el
colesterol, que mencionamos con anterioridad. Es una molécula de pequeño
tamaño, más pequeña que los fosfolípidos. Se suele colocar entre las colas de
los fosfolípidos. Tiene un grupo polar pequeño.
 |
| Colesterol en la membrana |
 |
| Colesterol en bicapa |
Los lípidos da una asimetría típica a las membranas. Las
hemimembranas externa e interna son distintas. Esta asimetría viene dada ya
desde su construcción. Uno de los motivos es que los lípidos forman un papel
importante en su unión con las proteínas. Algunas proteínas necesitan unos
grupos químicos para funcionar, que son aportados por los lípidos. Hay
proteínas que requieren lípidos. En cada hemimembrana, las proteínas se sitúan
de forma preferencial en una zona, en un lado y en una posición concreta.
Otro ejemplo de asimetría se da, por ejemplo, en la membrana
de los eritrocitos. En la cara exterior, encontramos fundamentalmente
fosfatidilcolina. En la interior hay otros fosfolípidos.
Hay un tipo de lípidos, los glucolípidos, que llevan un
árbol de azúcares en la cabeza polar. Normalmente llevan un único árbol de
azúcares, cuya composición es variable. El sentido de estos no está muy claro.
Se cree que actúan como contactos de reconocimiento. Hay una serie de
identificadores celulares, se sabe que actúan como receptores. Por ejemplo, en
el cólera, la bacteria se une solo a unos glucolípidos de las células
intestinales.
Proteínas de membrana.
Son un componente que le permite que las membranas adquieran
propiedades evolutivas y adaptativas: permiten el transporte, hace que la
membrana sea un buen sitio para que se lleven a cabo reacciones químicas, etc.
Se han estudiado mediante métodos de fraccionamiento
celular. Se ha visto que hay unas proteínas menos relacionadas con la membrana,
que se sueltan con facilidad y otras más relacionadas, que no se sueltan
fácilmente. Se clasifican en extrínsecas e integrales. Las integrales se unen
muy bien a la membrana. Las extrínsecas no suelen unirse con fuerza, están
ligeramente apoyadas sobre la bicapa lipídica. Para extraer las proteínas
integrales debemos destrozar la membrana.
 |
| Proteína de membrana |
Las técnicas de purificación son cada vez más sofisticadas y
evolucionadas y cada vez se conocen más y mejor las proteínas de membrana. Se
suele usar, para su estudio, la membrana del eritrocito.
Dentro de la membrana podemos encontrar varios tipos de
proteínas.
Por un lado están las proteínas transmembranales. Dentro de
estas podemos diferenciar dos tipos, las que atraviesan la membrana una vez y
las que la atraviesan varias veces. El sector que atraviesa la membrana suele
encontrarse en forma de hélice α (aunque en ocasiones también podemos encontrar
zonas del interior de la membrana en forma de lámina).
 |
| Proteínas integrales. |
 |
| Proteínas unidas a fosfolípidos |
 |
| Proteína unida a glúcido de un glucolípido |
 |
| Proteínas unidas a proteínas. |
 |
| Complejo protéico |
 |
| Proteínas de membrana unidas al citoesqueleto |
 |
| Proteínas de membrana |
 |
| Movimientos de proteínas |
Este tipo de experimentos se repitieron con otras
metodologías, consistentes en marcar con fluorocromos los determinantes
antigénicos. Posteriormente se decoloran los determinantes de una zona de la
célula, anulando la fluorescencia (usando normalmente una luz láser débil, que
al incidir sobre la zona agota la capacidad del fluorocromo de emitir luz). La
mancha sin color se mantiene un tiempo variable, pero acaba desapareciendo,
pues las proteínas de membrana van ocupando esa zona. Se puede medir el tiempo
que tardan las proteínas en ocupar la zona y hacer tablas, midiendo de este
modo la velocidad de la membrana. Mediante esta técnica se ha podido establecer
que la membrana más rápida es la de los fotorreceptores.
A la célula le interesa que exista una movilidad, pero esta
debe ser controlada. En las células se establecen dominios de membrana, zonas
de frontera en cuyos sitios se pueden mover las proteínas, pero de los cuales
estas no pueden salir.
Hay dos factores que modulan la rapidez con la que se
mueven. Por un lado, la fluidez de la bicapa. Por otro, el tamaño o peso
molecular de la proteína. Si tiene un peso molecular elevado, se mueve más
despacio.
En el caso de un organismo pluricelular, como ocurre por
ejemplo con las células epiteliales del intestino delgado, aparecerán células
con una polaridad elevada. En el caso concreto de las células epiteliales del
intestino delgado, estas presentan una zona con microvellosidades que mira a la
luz del tubo y una zona inferior que se encuentra en contacto con el tejido
conjuntivo y los vasos sanguíneos. Cada polo requiere un componente protéico
específico, las proteínas no se pueden mezclar (funcionaría mal y habría un
gasto excesivo de proteínas).
 |
| Dominios de membrana |
 |
| Dominio en Zónula Ocludens |
 |
| Proteínas de anclaje en dominios de membrana |
En cuanto a las glicoproteínas o glucoproteínas, estas se
encuentran ubicadas en las membranas, en la zona que mira hacia el espacio
extracelular o en las membranas interiores, mirando hacia al hueco de la
vesícula o del orgánulo en cuestión. Es decir, siempre miran al lado contrario
al citoplasma.
Los árboles de azúcares parecen relacionados con fenómenos
de reconocimiento celular y mecanismos de filtrado. Constituyen lo que se
denomina glicocálix. Y resulta muy variable, apreciándose grueso en algunas
células y más fino en otras.
No obstante, los árboles de azúcares son de mayor tamaño
cuando se encuentran anclados a los lípidos (glucolípidos) que cuando se
encuentran anclados a proteínas (glicoproteínas).
No hay comentarios:
Publicar un comentario