Características generales.

Además de las piezas básicas alimentadoras, dos o tres
piezas del pereion cambian de forma, se asemejan a las piezas bucales y se
caracterizan para la alimentación. Tiene un par de ojos compuestos. También dos
pares de antenas, las primeras antenas o antenas I y las segundas antenas o
antenas II. A las antenas I se les llama anténulas y a las antenas II antenas.
Son birrámeas. Tienen ciegos gástricos en el merodeo, son aminotélicos y tienen
celomiductos. Los gonoporos se abren a sitios diferentes, pero siempre cerca de
la separación entre pereion y pleon (en las últimas del primero o las primeros
del último). El desarrollo de los crustáceos, la segmentación del huevo, es
espiral obteniéndose de forma general larvas Nautilus.
Tiene solo tres pares de apéndices.
Encontramos unas cuarenta mil especies, marinas casi todas.
Algunas son dulceacuícolas y aun más raros, terrestres y viven en zonas
saturadas de vapor de agua.
Tegumento.
El tegumento posee, generalmente, una estructura mal
esclerotizada. Son tegumentos blandos. Cuando se endurecen lo hacen por
acúmulos de carbonato de calcio. No tiene capa de ceras. La capa de cemento
está mal esclerotizada, denominada epicutícula. La característica diferencial
está en la exocutícula. Esta tiene dos capas, cuando está calcificada. La
externa, denominada pigmentaria, es similar a la exocutícula de otros
artrópodos y se encuentra recubierta externamente por una capa calcificada, con
sales de calcio cristalizadas.
Nos encontramos siempre apodemas laminares abundantes y muy
desarrollados. Los tagmas son de dos tipos. El Cefalonia nomerísticos, es
decir, con un número de segmentos fijado: el acron y seis segmentos más. Y los
tagmas siguientes que son anomerísticos, tanto dentro de la superclase como
dentro de algunas clases.
Cabeza, tórax y abdomen.
Tiene dos pares de antenas birrámeas bien desarrolladas,
ojos compuestos, ocelos bien desarrollados (hasta ocho), con los que no podrán
ver figuras nítidas, pero detectan claridad y son capaces de fijar la posición
de focos luminosos, ver el origen de la luz. El labro está bien desarrollado.
Los ojos compuestos pueden ser pedunculados, que son más primitivos, o sésiles.
Los metámeros de pereion se denominan pereionoitos y los del
pleon pleonitos. Y los apéndices pereiópodos y aleonópodos (o pleiópodos) respectivamente. Dos
o tres pereiópodos se ponen al servicio de la boca y se les llama maxilípedos,
habándose de primeros, segundos y tercerso maxilípedos. Con cierta frecuencia
en el telson aparece un apéndice bifurcado denominado furca.
 |
| Anatomía externa de un crustáceo. |
Caparazón.
El caparazón es un repliegue del esqueleto cefálico hacia
atrás ara proteger una parte del cuerpo (es esa parte que se quita de la gamba
cuando se le arranca la cabeza). Se origina a partir del cefalón, del segundo
segmento maxilar. El caparazón posee tegumento y la hemolinfa recorre toda la
estructura.
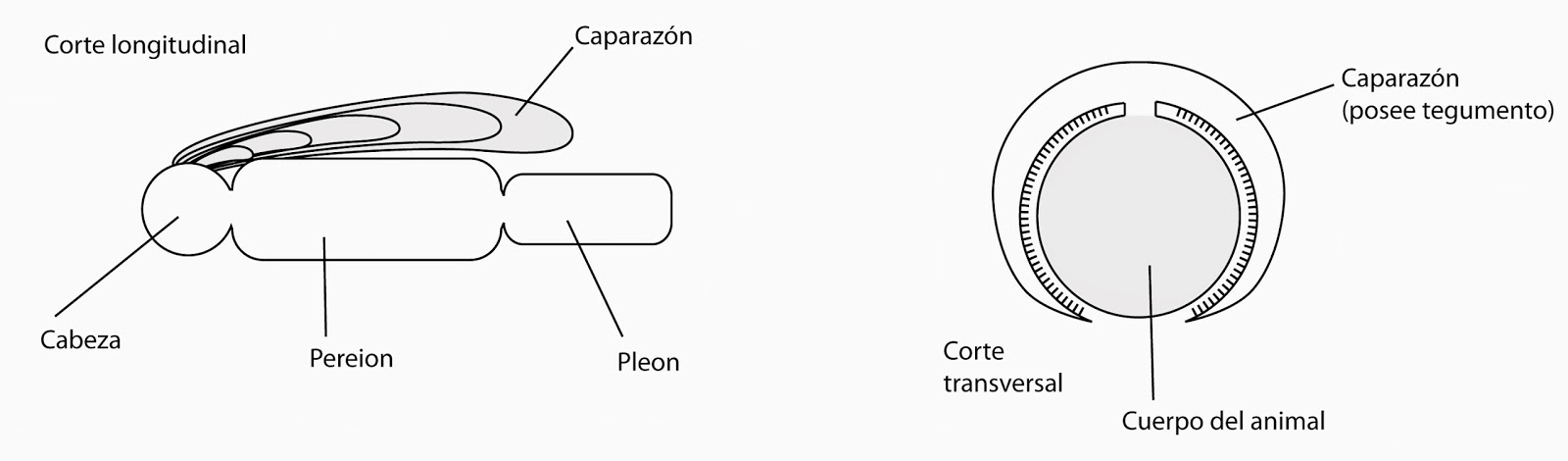 |
| Estructura del caparazón del crustáceo. |
El univalvo suele tapar el pereion, a veces también el
pleon. Se funden la parte dorsal con los pereionitos. Aunque se tapa el pleon,
nunca se funden con los pleionitos.
En los caparazones bivalvos el animal tiene un problema, ya
que el caparazón tapa al animal, pero también a los ojos y las antenas. Para no
hacer huecos para los ojos y las antenas, la zona se decolora, se hace
transparente (a modo de ventana). Para las antenas el caparazón tiene unas
escotaduras y las antenas quedan libres desde la base.
La función del caparazón es proteger al animal.
Secundariamente, la capa externa se ha endurecido, pero la interna se va
adelgazando y es bastante flexible y muy permeable. La cara interna del
caparazón es ideal para una respiración cutánea especializada. Esta
especialización aparece en casi todos los crustáceos. Además, también tiene
función de incubar los huevos. La hembra los deposita entre la cara interna del
caparazón y su cuerpo. Tiene un resalte que cierra parte de la cavidad y ahí se
desarrolla, al menos el embrión, pudiendo también tener lugar la fecundación.
Alimentación.
Pueden ser micrófagos o macrófagos. En los filtradores, los
apéndices tienden a ser planos y rodeados por franjas de pelos. Son como redes
que recogen el alimento. El movimiento de los apéndices lleva el filtrado hacia
la boca.
En los macrófagos encontramos predadores, fitófagos,
carroñeros, etc. El apéndice tiende a ser similar al de los insectos, alargado
y con un solo eje, acabando en quelas o subquelas destinadas a coger la presa o
el alimento.
Reproducción.
Son anfigámicos y ovíparos. La ovovivipiridad y viviparidad
no son frecuentes. Lo que si es frecuente es la partenogénesis. Los huevos son
protegidos frecuentemente por la madre. Lo hace de dos formas: cementarlos con
los apéndices o recogerlos y guardarlos en cavidades o sujetos con apéndices
sin fijación.
El desarrollo del huevo es holoblástico, desigual y espiral,
al menos en teoría (en realidad, el desarrollo espiral auténtico solo se
conserva en cirrípedos). Del huevo nace una larva nauplius, con desarrollo
anamórfico (tiene menos pares de apéndices que el adulto). Se transformara en
una larva metanauplius. Y después de dos o tres fases larvarias diferentes. La
forma juvenil se parece al adulto.
Clase Maxilipodos.
Podemos dividir los maxilípodos en dos grandes sublcases,
denominads Subclase Copepoda y Sublcase Cirripeda.
Subclase Copepda.
Los Copépodos suelen seer animales marinos. No tienen
caparazón ni ojos compuestos, solo ojos sencillos compuestos por tres o cuatro
ocelos. Perciben con ellos diferencias entre intensidad de luz. Las primeras
antenas o antenas I son mayores que las antenas II y son nadadoras. El
segemento I y el segmento II se unen en el cefalon. Posee apéndices birrameos
nadadores en el pereion.
La respiración es cutánea y el orificio genital se abre en
el primer segmento del pleon. Hay Copépodos de vida libre y parásitos.
Durante la reproducción, el macho agarra a la hembra con las
antenas y coloca el espermatóforo en los orificios de la hembra. Es
espermatóforo tiene empaquetados los espermatozoides. La hembra alimenta los
huevos, forma masas cementadas.
 |
| Esquema de un espermatóforo de un copépodo. |
Los huevos eclosionan dando lugar a una larva tipo nauplius,
apareciendo posteriormente una metanauplius. Esta no va dar lugar directamente
al juvenil, sino que pasará por otra fase larvaria, el copepodito.
Forman grandes biomasas. Son alimento de otros seres vivos.
En casi todos no encontramos con fenómenos de migración vertical, es decir, los
individuos hacen viajes hacia arriba y abajo del las masas de agua. ascienden
por la noche y descienden durante el día. Este fenómeno sucede en los que viven
cerca de la superficie y también en los de las profundidades, aunque no
perciban luz.
 |
| Copépodo |
Subclase Cirripeda.
Los Cirrípedos son artrópodos extraños. No se parecen a
crustáceos, ni siquiera parecen artrópodos. En un primer momento, se creía que
eran moluscos y no se descubrió su origen artrópodo hasta que se vio el
desarrollo postembrionario.
 |
| Cirrípedo. |
Son un grupo pequeño, de unas novecientas especies. Y son
sedentarios o parásitos, tanto endoparásitos como exoparásitos. Presentan
caparazón bivalvo, la cabeza va a ser relativamente grande, tienen todos ellos
anténulas que poseen glándulas cimentantes. Pueden sujetarse por medio de ellas
al sustrato.
Las antenas II están muy reducidas. Durante el desarrollo
presentan ojos compuestos que degeneran y no llegan al estado adulto. El
pereion tiene seis pares de apéndices birrámeos. La larva nauplius tiene
siempre antenas a modo de curnos. Muda seis veces y origina siempre una larva
denominada larva cipris que tiene solo seis pares de pereiópodos y un caparazón
bivalvo, sujeto por un solo segmento. Esa larva tiene ojos compuestos y sufre
una metamorfosis muy intensa.
Solo hay dos órdenes de Cirrípedos, el Orden Thoracica, que
presenta aspecto de crustáceo, y el Orden Rhizocephala, animales muy extraños.
Los Cirrípedos del orden Thoracica son los percebes. Las
antenas I de la larva cipris tienen glándulas cimentantes. La larva busca un
sustrato, se paga por la antena y comienza a metamorfosearse. La antena I y la
región cefálica próxima se engrosan, crecen y forman el pie del percebe. El
resto del animal se coloca con el dorso sobre este. Pierde los ojos compuestos. Las valvas se
calcifica, no de forma continua, sino selectivamente. Se forman placas, estrías
de crecimiento. La estructura interna es la típica de un crustaceo modificada,
el sistema nervioso está muy simplificado, poseen un ovario y testículos, es
decir, son hermafroditas, pero requieren fecundación cruzada, presentando el
pene más largo del reino animal.
Los del orden Rhizocephala son siempre endoparásitos de
otros artrópodos. Parasitan cangrejos, por ejemplo. Algunos penetran en el
interior de las hembras por un poro genital y atraen a los machos, que se
quedan en forma de larva. Se acercan al poro genital y se ponene los huevos,
que salen al agua. de ahí nace la larva nauplius, que derivará en una
metanauplius y una cipris. Si son machos, el desarrollo se para ahí. Llega un
momento crítico en el que las hembras necesitan encontrar un animal que
parasitar, un cangrejo, para poder sobrevivir. Si lo encuentra, se fija sobre
la base de un pelo den cangrejo. Debe fijarse a la base. Cementa una sola
anténula. Y sufre metamorfosis, pasa a la forma de larva cipris fijada. Pierde
el pereion y todos los apéndices, cambia la forma de su Cefalonia que se hace
plano y pierde los ojos compuestos. Queda reducido a apenas una masa de
células. Se rodea de una cutícula externa, pierde el caparazón bivalvo y se
convierte en la larva kentógena. Se produce una segunda envuelta interna, con
un pincho, una aguja hueca. Esta larva aumenta la presión de la masa, la aguja
se evagina, se clava en la base del pelo y llega al interior del animal. Todo
el contenido celular pasa por la aguja al interior del cangrejo, a modo de masa
indiferenciada. Si el animal se ha pegado al pelo de un cangrejo que iba a
mudar, no le dará a parasitarlo, al mudar se perderá con el caparazón. Y si se
une a una anténula que no esté fijada, vierte el contenido al exterior en lugar
del interior y también morirá. Pero si la masa celular logra entrar al interior
del cangrejo, mediante movimientos ameboides se colocará en el tubo digestivo,
se extiende, forma una especie de raíces y se instaura sin matar al cangrejo.
Pero producirá castración parasitaria y cambiará su morfología externa, dando
a la hembra caracteres masculinos y viceversa. El parásito forma un ovario
durante la época reproductora, crece, comprime la membrana articular de esa
zona la rompe, abriendo al exterior el
poro genital (todo lo que podríamos ver es un amasijo de células en el interior
del cangrejo).
Clase Malacostracea.
Características de los Malacrstráceos.
Son los crustáceos más evolucionados. Todos los taímas son
nomerísticos. El pereion y el pleon tienen el mismo número de segmentos
siempre, ocho y seis respectivamente. Los gonoporos se encuentran en el octavo
pereionito en machos y en el sexto pereionito en hembras.
Los pleiópodos están bien formados y son monorrámeos, como
los de los insectos, salvo aquellos que están al servicio de la boca. los
pleiópodos están mal formados y son birrámeos. El cuerpo acaba en un telson sin
furca y de forma bastante común aparece un abanico caudal, formado por el
telson, laminar y por un par de apéndices birrámeos del sexto pleonito. Los
ojos son compuestos en general y pedunculados, apareciendo sésiles en los más
evolucionados.
 |
| Anatomía general de un Malacostráceo. |
 |
| Esquema del tubo digestivo del un malacostráceo. |
Clasificación de los Malacostraceos.
En los Malacrostaceos encontramos dos superórdenes, los
Paraceridae y el Euceridae. Analizaremos los órdenes más importantes de cada
uno de ellos.
Superorden Paraceridae.
Son los malacrostaceos más evolucionados. No suelen
presentar caparazón y los pocos que lo presentan, no les cubre todo el pereion.
En las hembras una parte de los apéndices aparecen aplicados contra la cara
ventral y son planos, denominándose hoosteitos. Estos dejan una cavidad
incubadora. Del huevo nacen siempre juveniles.
Los ojos son pedunculados y sésiles en los más
evolucionados. Esa presencia de ojos sésiles y juveniles son caracteres
evolutivos muy superiores a los que van a presentar los cangrejos.
Se diferencian dos órdenes importantes (hay muchos más). Por
un lado, el orden Anphipoda, que son las pulgas de arena, con cuerpo arqueado y
los tres últimos pereiópodos mas largas, sirviéndoles para saltar.
 |
| Anfípodo. |
En general todos ellos son bentónicos. Suelen ser marinos,
algunos de agua dulce e incluso semiterrestres.
El orden Isopoda no tiene
caparazón. Los apéndices respiratorios de los crustaceos,
generalmente, se
encuentran en el pereion, pero en este grupo se encuentran en el pleon. Los
Isópodos son terrestres o semiterrestres, apareciendo especies marinas. Viven
siempre en zonas saturadas de vapor de agua. Pero existen especies que viven,
incluso, en el desierto.
 |
| Isópodos. |
Superorden Euceridae.
Son los cangrejos verdaderos. Se trata de crustaceos más
primitivos que los anteriores. Existen unas nuevemil especies. Poseen ojos
compuestos pedunculados. Todos presentan caparazón. El caarazón está fundido
con los pereionitos. Solo vamos a analizar dos órdenes.
Por un lado, el Orden Eupheusiceae son un grupo de animales
que viven mar adentro. Hay unas cien especies que pueden formar biomasas
enormes. Todos ellos son filtradores. A diferencia de los decápdos (el otro
orden que veremos) todos sus pereiópodos son ambulatorios, ninguno queda al
servicio de la boca. las bránqueas quedan siempre al exterior y el caparazón
desciende poco por los lados, apenas los tapa.
En cuanto al Orden Deapoda, el caparazón tapa las branquias.
Los tres primeros apéndices del pereion están modificados en forma de
maxilípodos, quedando cinco pares de pereiópodos (de ahí su nombre de
decápodos).
 |
| Decápodos: langosta (natatia) |
 |
| Decápodos: cangrejo (reptantia) |
Los Anomura tienen la cola de forma extraña, siendo la mayor parte de los cangrejos ermitaños. En estos, la
parte del pleon será blanda. La característica del grupo es que los dos últimos pares de pleiópodos no son marchadores, son muy pequeños y están doblados hacia arriba. Esto permite reconocer a los libres. Por úlimo los Brachyura (que significa literalmente cola pequeña) no se ve aparentemente el pleon, está plegado por debajo del cefalotórax.
























