ARN polimerasas.
El ADN se traduce para formar ARN, bien ARNm, ARNt o ARNr. A
partir de estos tres tipos de ARN se fabrican las proteínas (como hemos visto en entradas anteriores).
Las reacciones que catalizan las reacciones de fabricación
del ARN son las ARN-polimerasas ADN-dependientes. En procariotas como
Escherichia coli, solo hay un tipo de ARN-polimerasa.
En eucariotas habrá tres (que se nombran con números
romanos). La ARN-polimerasa I sintetiza los ARNr 28s, 18s y 5,8s. La
ARN-polimerasa II sintetiza el ARNm. La ARN plimerasa III sintetiza ARNt y ARNr
5s.
Estudiaremos la ARN-pol de Escherichia coli. Se trata de un
enzima de cinco subunidades: dos subunidades α2,
una subunidad β, una subunidad β’
y una subunidad θ. Las dos subunidades α2 y las β y β’
constituyen el núcleo del enzima. Y tienen distintas funciones.
 |
| Funciones de la ARN polimerasa |
Por un lado están las funciones polimerásicas. En esencia es
paralela o equivalente a la de la ADN plimerasa, formando ARN a partir de las
correspondientes ATP, GTP, CTP y UTP:
Necesita una molécula de ADN patrón para copiar la
secuencia. Pero no necesita cebador. Y necesita que los cuatro sustratos estén
a la vez, es decir, necesita que estén presentes ATP, CTP, GTP y UTP.
El pirofosfato es inmediatamente hidrolizado por la
pirofosfatasa (como ocurre en la replicación del ADN), ara asegurar que el
equilibrio se desplaza hacia la derecha:

El patrón es el ADN. Pero este tiene dos hebras, una en dirección 5’→3’ y la otra en dirección 3’→5’.
La hebra en dirección 5’→3’ es la hebra
antisentido y la hebra en dirección 3’→5’ es la hebra molde o
con sentido.
Ahora bien, cuál de las dos hebras se va a transcribir y
cual es la que no se transcribe depende, varía (debemos tener en cuenta que es
solo cuestión de cambiar de dirección, no hace falta cambiar de hebra).
El ARN que se forma debe ser complementario y antiparalelo a
la hebra molde, pero donde hay A ahora se enfrentará a una U en vez de a una T.
Se conocen casos en los que se transcriben las dos hebras.
Por ejemplo, en algunos plásmidos de bacterias. La dirección de síntesis es
siempre la dirección 5’→3’ y la dirección de
lectura es la de 3’→5’. Y el crecimiento en la
dirección 5’→3’ es idéntico a las ADN polimerasas.
 |
| Reacción de la ARN polimerasa |
Transcripción.
El proceso de transcripción suele dividirse en tres etapas:
iniciación, elongación y terminación.
Para la iniciación hay una secuencia de alrededor de
cuarenta pares de bases en el ADN que marcan dónde tiene que empezar. Dentro de
esta secuencia, hay dos que le dan la polaridad. Esa secuencia es reconocida
por la ARN polimersa y hace que se desarrollen el ARN a unas 17 pares de bases
de distancia. La ARN-pol recorriendo el ADN por el surco profundo hasta que
reconoce esta secuencia, estereosespecíficamente y comienza separándose de ella
unas 17 pares de bases.
Aunque el ADN podría leerse en los dos sentidos, la
secuencia de iniciación estará colocada de forma que el enzima, al encontrarla,
ya quebrará colocado para leer en una dirección, hacia uno de los lados.
La primera base que se suele poner es una purina, bien A o
G:
PPPG --- NTP + NTP → NTP-NMP
Se sintetizan unos cuantos núcleotidos, que marcan como
colocarlos. En esto estaría implicada la proteína θ
de la polimerasa, que ahora se separa y a partir de aquí, solo irá el núcleo
del enzima.
A partir de ahora tiene lugar la segunda fase, la elongación
de la cadena.
 |
| Elongación de la cadena |
Se forma una especie de burbuja que avanza por el ADN y de
ahí su nombre, burbuja de transcripción. En un momento dado, solo estarán los
últimos 12 nucleótidos emparejados con el ADN, formando un híbrido ADN-ARN. El
resto van separándose, es decir, los que se han fabricado primero se van
separando.
Se sintetizarán alrededor de 50 nucleótidos por segundo, es
decir, es más lenta que las ADN polimersas. En este caso, no va a haber un
proceso de corrección. Comete muchos más errores, alrededor de un factor de
10-6 errores (un error por cada millón de nucleótidos). Pero en este caso los
errores son menos trascendentales. No se produce una mutación, el error es
temprano y no se hereda, por eso no hay tanta necesidad de corrección.
 |
| Secuencia de terminación |
La última fase es la terminación. Hay unas secuencias de
terminación. Cuando la ARN-pol se encuentra con una secuencia muy rica en G-C,
se autocompletaría. Se forma una especie de bucle que finaliza la
transcripción. Tras la secuencia rica en G-C, hay varios uracilos seguidos. Es
la señal para que la ARN-pol se separe, se desprenda.
Algunos organismos necesitan proteínas adicionales que
ayudan a la separación. En Escherichia coli se encargan las proteínas β, proteínas capa y algunas otras.
En eucariotas, pero también en bacterias y virus es
frecuente que un grupo de distintos genes compartan la misma secuencia
iniciadora y finalizadota, con lo que un ARN puede contener la información de
distintos genes. Después de traducirse, o durante la traducción, se cortarán.
Se habla de poligenes o genes policistrónicos.
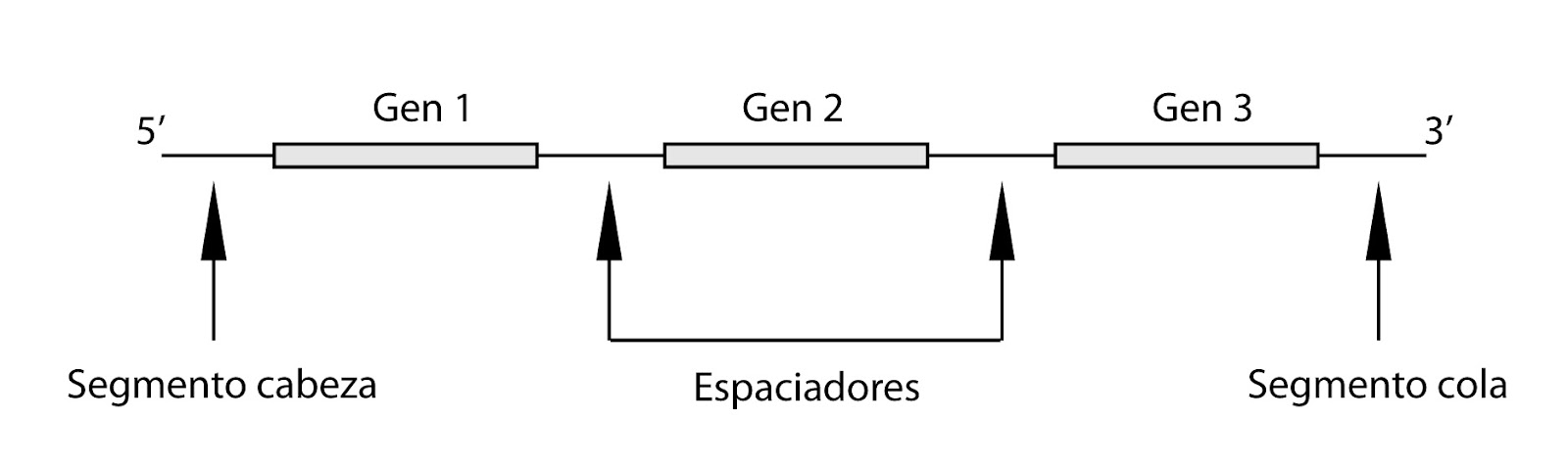 |
| Genes policistronicos |
Los genes policistrónicos suelen tener una relación
funcional, por ejemplo, pertenecer a una misma ruta metabólica.
En eucariotas, normalmente, los ARN son poligénicos o
policistrónicos.
Procesamientos postranscripcionales.
Muchos ARN pueden pasar a realizar su función, a ser
traducidos. Pero esto no ocurre siempre. Suelen ser necesarios algunos
procesamientos postranscripcionales. Estos procesamientos son la mayor
diferencia entre eucariotas y procariotas a la hora de fabricar ARNm.
En los procariotas el ARNm prácticamente no tiene que
procesarse. Pasa directamente a ser traducido. Es muy frecuente que los ARNm se
estén traduciendo aún cuando, aun sin haber terminado, sin ser trasncrito del
todo, ya está siendo traducido. Este proceso se ve favorecido porque en
procariotas no hay membrana nuclear. Y tras transcribirse y traducirse, se
degrada (puede incluso comenzar su degradación cuando el proceso de traducción
no ha acabado del todo, destruyéndose por la zona que ya ha sido traducida).
En el caso de eucariotas esto no puede tener lugar, ya que
los distintos procesos se llevan a cabo en lugares diferentes. El ARNm se
transcribe en el núcleo, pero es en el citoplasma, fuera del núcleo, donde
tendrá lugar la traducción y fabricación del proteínas. Aldemás, antes de salir
del núcleo, el aRNm suele tener un proceso de maduración, el procesamiento
postranscripcional (solo hay casos puntuales en los que no se necesita este
procesamiento, como en el ARN que codifica a las lisozimas del espermatozoide).
En eucariotas el ARNm es transcrito por al ARN-pol II. Serán
genes estructurales, que darán lugar a poliméptidos, aunque también se usa ese
enzima para fabricar algunos ARNr. Los genes estructurales son los genes
determinantes del ADN.
Maduración del ARNm.
Veamos el proceso de maduración del ARNm. Al ARN que se
fabrica en el núcleo se le denomina ARN heteronuclear, ARNhn.
El primer cambio es la modificación de los extremos 5’ y 3’
del ARNhn, haciéndolo más resistente a la degradación. En el extremo 5’ se le
adicciona un residuo de trimetilguanina, unido por un puente o enlace
trifosfato. Esto puede llevarse a cabo antes de que el ARN se acabe de
traducir. Se habla de caperuza o casquete, 5’cop.
Por otro lado, posteriormente, en el extremo 3’ se le añade
una cola de ácido poliadenílico (poli A). Esta reacción es catalizada por la
poli-A-polimerasa (añade entre 50 y 250 adeninas consecutivas).
 |
| Maduración del ARN mensajero |
En algunos casos aislados, en procariotas, tiene lugar una
especie de splicing o empalme, en el que se eliminan los segmentos de ARN no
codificantes o ARN intrónico. Se da, por o tanto, en genes fragmentados o
discontinuos. Aunque se trata de un proceso poco frecuente en porcariotas, es
muy frecuente en eucariotas.
Genes fragmentados o discontinuos.
Un gen contínuo es aquel en el que el gen codifica una
secuencia de aminoácidos continua. En el caso de genes fragmentados lo que
ocurre es que la secuencia de nucleótidos, que codifican para los aminoácidos,
no están de forma continua. Hay regiones de los genes que no codifican para la
proteína. Y se denominan intrones. Las zonas que codifican para la proteína se
denominan exones.
Tras la transcripción se genera un ARN que contine intrones
y exones. Se le añade el 5’-cop y se poliadeniza. Ahora deben ser eliminados
los intrones, por un proceso denominado splicing. Es llevado a cabo por un
complejo denominado RNPsn (small nuclear), que tiene unos pequeños trozos de
ARN, de unos 300 nucleótidos denominados ARNsn (ARN small nuclear). Este ARNsn
se empareja con el ARN intrónico por los extremos, para unirlo por los extremos
formando una pequeña doble hélice, por donde se realizará el corte para
eliminar el intrón. Debe ser un proceso extremadamente preciso, ya que en
contrario, se estropearía el ARN.
 |
| Spilicing |
Al final del proceso obtenemos el ARN maduro, que puede
pasar a ser traducido. Ya no tiene ARN intrónico.
Ahora se debe unir por la cola de poliadeninas (poli-A) a
unas proteínas que lo saquen del núcleo al citoplasma, donde será traducido.
En muchas especies, el splicing puede producirse de varias
formas, pueden tener lugar de diferentes modos para un mismo ARN original,
pudiendo por lo tanto obtenerse varios tipos de ARN maduro.
No hay comentarios:
Publicar un comentario