 |
| Células: marcaje de microfilamentos |
Son el tipo de filamento más pequeño. Está constituido por
subunidades de actina. En las células musculares, sobre todo en el músculo
estriado, hay una disposición muy ordenada del citoesqueleto, sobre todo de las
fibras de actina y de miosina, lo que hace que se constituya un citoesqueleto
muy fuerte (que no se pierde en el proceso de fijación) y relacionado con las
reacciones que permiten el movimiento.
Los microfilamentos forman una trama que es muy frecuente en
las células. Constituye la estructura de esterocilios, microvellosidades, etc.
Como indicábamos, están constituidos por actina. Es una
proteína con un elevadísimo grado de conservación evolutiva. Aparece de dos
formas en la célula, con la forma G y con la forma F. La G es la forma en la
que se encuentra como reservorio. La F es la forma en la que se encuentra al
formar los microtúbulos. La forma F (de filamentosa) es la polimerización de la
forma G (globular). Y la actina está continuamente variando estre estos dos
estados.
La forma G suele venir asociada a una molécula de ATP. Se
puede inducir el paso de actina G a actina F “in Vitro” y conseguimos que el
líquido en el que teníamos aislada la actina G se vuelva viscoso. El ATP se
degrada en el proceso, pero se sabe que la polimerización en global no necesita
consumir ATP. Esto se demuestra porque, si añadimos ADP a la forma G, sigue
produciéndose polimerización aunque del ADP no le sea posible obtener energía.
La asociación con el ATP se debe a otros fenómenos, como el recambio de la
actina globular.
 |
| Modelo de filamento de actina. |
La asociación de los microtúbulos con otros elementos es muy
variad. No todos los fenómenos mediados por los filamentos de actina se pueden
explicar por procesos o fenómenos de polimerización y despolimerización. Se ha
visto que hay relación de actina con otras proteínas. Las proteínas que se
encargan de inducir la formación de tramas son denominadas gelificantes. Se
asocian entre ellas y la más conocida es la filamina, que se estructura
formando dímeros. Dan mayor consistencia a los microfilamentos y permiten
cruzamientos. También hacen las veces de membrana, se se hace una presión
fuerte, es dúctil y se deforma, volviendo por si solo a la posición original. Pero
si se hace presión continuiada y suave, se deforma perdiendo la elasticidad.
Otras proteínas asociadas a los microfilamentos son las que
se encargan de estabilizarlo, uniéndose a el y manteniendo su estabilidad. El
ejemplo más típico es la tropomiosina, relacionada con la contracción muscular.
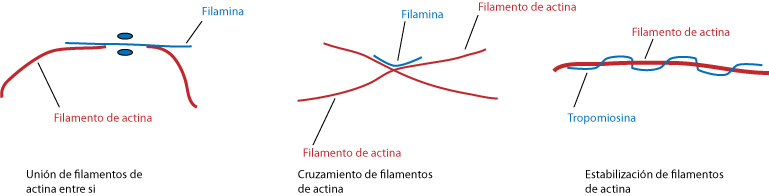 |
| Proteínas asociadas a los microfilamentos. |
Otro tipo de proteínas son las fragmentadotas. Inducen una
despolimerización, rompiendo la actina F y transformándola en actina G.
También existen proteínas antipolimerizantes, que se asocian
a la actina G impidiendo que se polimerice y pase a formar actina F. Esto es
importante para tener reservorios de actina, de forma que pueda fabricarse y
acumularse sin que polimerice.
Hay proteínas que median la unión con la membrana
plasmática. Un ejemplo es la espectrina, que actúa como elemento intermediario.
Otras inducen el desplazamiento de los filamentos. Por
ejemplo la miosina mueve unos filamentos respecto a otros (contracción
muscular, por ejemplo).
También las hay que inducen el desplazamiento de vesículas
sobre la actina. Un ejemplo es la minimisina. Se postula la existencia de
proteínas de tipo capuchón cuya función sería estabilizar el extremo (+) del
filamento, aunque no están completamente identificadas.
Los movimientos de los filamentos de actina son fácilmente
observables en las células musculares gracias a su ordenación. Pero en otros
tipos celulares forman una maraña en la que es difícil apreciear este tipo de
procesos.
Funciones de los microfilamentos.
Podemos agrupar las funciones en dos grandes tipos, las que
tienen que ver con la contracción celular y la que tiene que ver con la corteza
citoplasmática. En cuanto a la corteza citoplasmática, podemos dividir en tres
tipos, mantenimiento de la disposición de la membrana, proyección de la
membrana plasmática e intrusión de la membrana plasmática.
 |
| Estructura de actina y miosina en las células musculares. |
 |
| Microfilamentos y contracción o movimientos celulares. |
Este modelo permite explicar sistemas que no están
relacionados con la contracción, sino con el desplazamiento de estructuras
sobre el filamento, similar al que vimos en los microtúbulos. Este proceso se
ha visto en algas plinucleadas, en las que se aprecia movimientos del citoplasma
alrededor de una vacuola. Los filamentos de actina están medianamente ordenados
en estas células y marcan las direcciones de las corrientes del citoplasma. Y
el proceso está mediado por la minimiosina.
 |
| Movimientos por el citoplasma mediados por microfilamentos |
Como puede verse en los esquemas, la cabeza de la
minimiosina tiene afinidad por la actina. En cambio la cola tiene afinidad bien
por ella misma, bien por la unión a otra minimiosina mediada por otra proteína,
bien por la membrana plasmática.
Pasemos ahora a hablar del mantenimiento de la disposición
de la membrana (correspondiente a la corteza plasmática). La estructura más
estudiada es la del eritrocito de mamíferos, con su morfologíaa de disco
bicóncavo. Su morfologíaa varía en algunos sitios, dependiendo del grosor del
capilar por el que se mueva. Su morfología no es la que tendría de forma
natural, ya que debería ser estérico y se debe, por lo tanto, a la acción de
los microfilamentos.

Es importante la espectrina, que está relacionada con la
actina. Existen otras proteínas que también se relacionan directa o
indirectamente de la actina. Todo el conjunto se forma durante el proceso de
maduración y ayuda a mantener la estructura.
 |
| Filamentos de actina en uniones adherentes. |
 |
| Estructura de los microfilamentos en microvellosidades. |
Los filamentos que se unen a la membrana plasmática y a los
filamentos de actina son moléculas de minimiosina. Se pensaban que servían solo
de anclajes, pero hoy se postula que hay más una renovación continua de la
membrana de las microvellosidades. Es decir, se recicla la membrana. En las
microvellosidades intestinas se va desprendiendo al exterior, se digiere y de
esta forma hay una renovación contínua. Y son las microvellosidades las que
provocan estos movimientos de la membrana. Del complejo de material electrondenso
de la zona apical no se tiene demasiada información.
 |
| Micrografía electrónica de microvellosidades. Se aprecian microfilamentos. |
 |
| Extrusión de membrana mediado por microfilamentos. |
 |
| Cabeza de espermatozoide con acrosoma |
En los espermatozoides también encontramos procesos de
extrusión de membrana. Al legar al óvulo, en algunas especies, al
espermatozoide le sale una prolongación “a modo de rompa” que llega al óvulo y
que se denomina filamento acrosómico. En la cabeza, antes de llegar al núcleo,
encontramos actina en forma globular, bloqueada or proteínas. Al llegar al
óvulo tiene lugar una variación del pH, que hace que se libere actina G de las
proteínas que estaban bloqueando a la actina. Se polimeriza como actina F
produciéndose el alargamiento (formación del filamento acrosómico).
En la fagocitosis hay una polimerización de filamentos de
actina. En fenómenos de desplazamento por espinas o filopodios también. Se da
una polimerización a modo de caden de tanque. Para comprobar el mecanismo se
han realizado estudios con actina G fluorescente y anulando la fluorescencia en
una zona mediante un láser. Puede comprobarse como esa zona sin fluorescencia
se va desplazando.
 |
| Movimiento de membrana y microtúbulos: formación de filópodos |
No hay comentarios:
Publicar un comentario