La variabilidad genética no surgió repentinamente. Todo
proviene de un grupo de células procariotas iniciales. La evolución se produjo
porque el material genético se ha podido ir modificando de generación en
generación. Hasta ahora, hemos estudiado la herencia como un proceso que
mantiene intacta la información, tanto en la replicación y reparación del ADN
como en la mitosis. Pero hay procesos que introducen variabilidad. En las
células eucariotas estos procesos son, básicamente, la mutación y la reproducción
sexual. En procariotas existen métodos distintos, ya que además de la mutación
encontraremos la conjugación o la transducción.
La mutación.
Entendemos por mutación cualquier cambio en el material
genético que, perfilado a nivel bioquímico, es cualquier cambio en secuencia,
orden o número de nucleótidos. Una vez se ha producido, este nuevo gen mutado
sigue utilizando los mismos mecanismos de la reproducción.
Tipos de mutaciones.
Podemos clasificar las mutaciones en distintos tipos.
Por un lado tenemos las aberraciones cromosómicas o
mutaciones de gran magnitud, que implican cambios como la variación en el
número de cromosomas (individuos 3n en vez de 2n, o con tres cromosomas de uno
de los tipos, por ejemplo) y alteraciones estructurales, visibles con el
microscopio óptico, como grandes delecciones, duplicaciones o traslocaciones,
por ejemplo.
Por otro lado tenemos las mutaciones génicas, que afectan a
un gen o a un carácter concreto. Puede decirse que afectan a un gen o cistrón.
Las mutaciones son errores y como tales son poco frecuentes.
Se llama tasa de mutación a la probabilidad de que tenga lugar una mutación
concreta en una célula o un individuo. Esta tasa ronda los 10-5. Por ejemplo,
la probabilidad de la hemofilia es de 3.10-5.
Las mutaciones son espontáneas, pero las tasas pueden
aumentarse por distintos factores o agentes mutagénicos. Por ejemplo, la
radiación ultravioleta, así como rayos α, β
o γ, que modifican las bases provocando más errores aunque
existan correcciones específicas.
Si realizamos una gráfica con las mutaciones según la dosis,
observamos que obtenemos una línea recta, en la cual se ve que el porcentaje de
mutación aumenta linealmente al aumentar la dosis de radiación.
 |
| Relación radiación/mutaciones |
También existen los mutágenos químicos, que tienen una
especificidad en su actuación. Tienen un efecto más retardado, pudiendo no
actuar sobre la primera generación y actuar después de la segunda, o después de
varios ciclos.
Existen otros mutágenos físicos, como los ultrasonidos, o
incluso los cambios bruscos de temperatura.
Las mutaciones son errores, es decir, son accidentales y al
azar. Son accidentes del sistema Bioquímica, porque derivan de que los sistemas
de reparación hayan funcionado mal. Por otro lado, recalcan el factor del azar,
porque nunca es predecible en una especie qué individuo puede presentar una
determinada mutación aunque sepamos el porcentaje de individuos que van a
presentar mutaciones.
Otro aspecto es que las mutaciones se presentan
independientemente de que sean beneficiosas, perjudiciales o indiferentes,
siendo lo segundo, es decir, que sean perjudiciales, lo más frecuente. Las
mutaciones no tienen porque mejorar la especie.
Por ejemplo, si cultivamos una bacteria sensible a un
antibiótico en presencia del antibiótico, siempre encontraremos alguna colonia
que sobrevive. Parece lógico pensar que el medio indujo la mutación. Pero se
trataría de un error, la mutación ya estaba presente entre las bacterias
originales, pero no se había manifestado.
Bases moleculares de la mutación.
El elemento más simple que podemos identificar en el ADN son
un par de bases. Son la unidad mutacional o mutona.
Las mutaciones se dividen en dos grupos, las sustituciones
de pares de bases y las alteraciones de corrimiento en la pauta de lectura.
Sustitución de pares de bases.
Las sustituciones en los pares de bases pueden dividirse en
dos grandes tipos, transiciones y transversiones.
Las transiciones son aquellas en las que se sustituye una
base púrica por otra base púrica o una base pirimidínica por otra base
pirimidínica. Es decir:
Las tranversiones son lo contrario, donde había una base
púrica ahora habrá una pirimidínica y viceversa:
Las transiciones son las mejor conocidas. Un ejemplo
espontáneo tiene lugar en la replicación por tautomería de las bases. Las
bases, por un desplazamiento de un hidrógeno dentro de la misma molécula, puede
pasar de las formas normales a otras menos frecuentes llamados tautómeros, que
son menos comunes (10-4).
Veamos como afecta a la adenina. Se produce un cambio de un
grupo amino (-NH2) a un grupo imino (=NH): -NH2 →
=NH
 |
| Transición de la adenina |
Ahora la adenina modificada (A*) se enfrentaría a la
citosina en lugar de hacerlo a la timina.
En el caso de la timina es un cambio de un grupo ceto (=O) a
un grupo enol (-OH): =O → -OH
 |
| Transición de la timina |
Los tautómeros evaden muy bien los sistemas de reparación
del ADN. Se producen espontaneamente y en la replicación. Tras la replicación
del ADN, la mutación quedaría instaurada.
 |
| Evasión de la reparación de los tautómeros |
Hay factores que modifican las bases. Estos cambios pueden
tener lugar en la replicación o fuera de la replicación. Un ejemolo es la
acción del HNO2, que actúa fuera de la repliación y que actúa sobre el -NH2,
provocando que sea sustituido por un radical –OH. Un ejemplo es sua actuación
sobre la adenina, atacando a su -NH2 y transformándola en una base no común
denominada palmatina. Sobre la citosina actuará transformándola en uracilo.
 |
| Transformación de citosina en uracilo |
Si el sistema de reparación no detecta el cambio de citosina
por uracilo, este se emparejará con la adenina. Por lo tanto se producirá un
cambio de un par C-G por un par T-A.
Aunque esta reacción está promovida por el HNO2, también
puede tener lugar expontaneamente dentro del ADN.
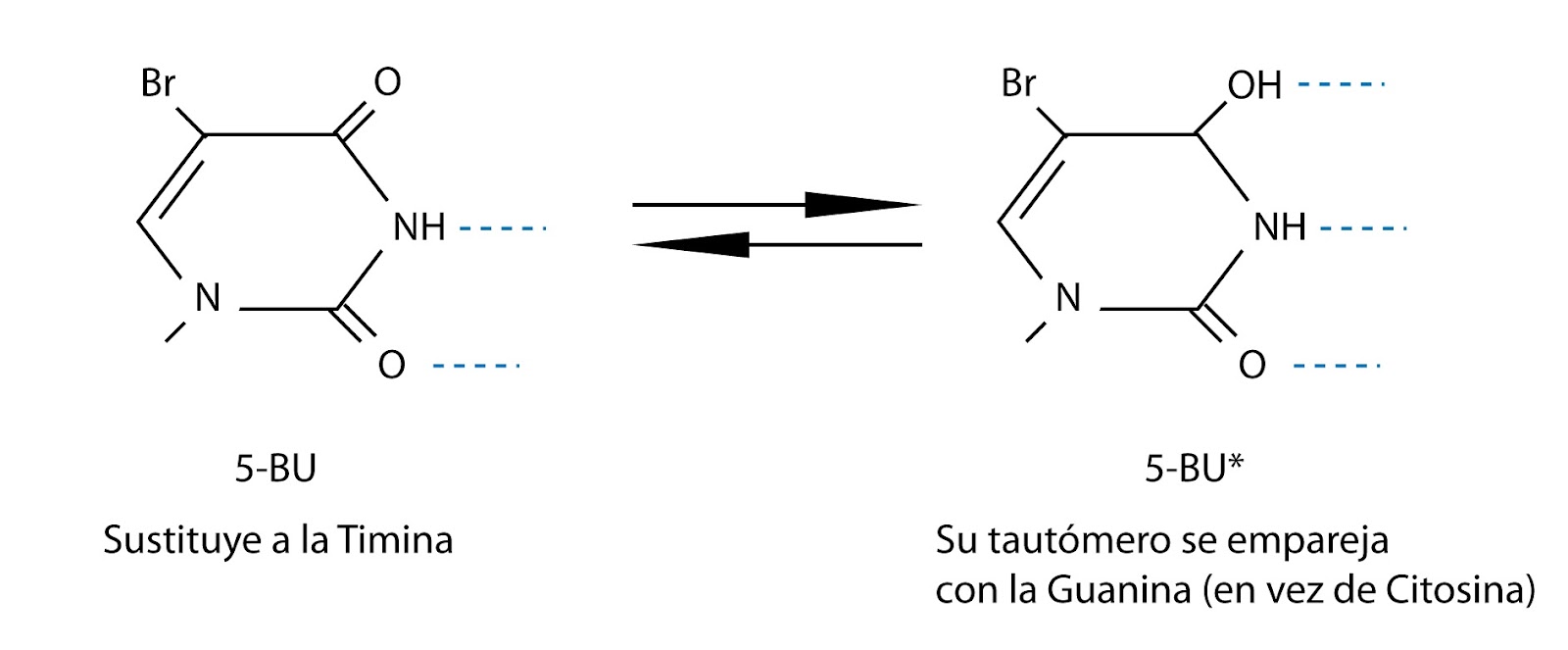
Hay compuestos mutágenos que actúan en la replicación. Por
ejemplo, sustancias que son parecidas a las bases. Una de ellas es el
5-bromouracilo. Esta base tiene mayor frecuencia de tautomerización, cambiando
a su forma tautómera.
5-BU → 5-BU*
el 5-BU es el
5-bromouracilo es su forma normal también denominada forma ceto, mientras que
el 5-BU* es su forma tautómera o forma enol.
 |
| Transformaciones del 5-bromouracilo |
El 5-bromouraciolo sustituye a la timina, pero su tautómero
funciona como la citosina. Es decir, pasa de combinarse con emparejarse con
adenina a emparejarse con guanina.
Veamos como tiene lugar la transición cuando el
5-bromouracilo sufre la tautomerización:
 |
| Transición con tautomerización del 5-bromouracilo |
Como vemos, tiene un efecto retardado. Actúa, al menos, en
la tercera replicación.
Y puede tener lugar la reacción contraria:
 |
| Reacción opuesta a la tautomerización del 5-bromouracilo |
El otro tipo de cambios posibles son las transversiones. No
se conocen muy bien. Pueden ser que en el ADN, durante la replicación, haya un
hueco, es decir, falte una base. Enfrente de ese molde vacío, se colocará una
base cualquiera. Esto es relativamente frecuente, sobre todo en el caso de
purinas, hablándose de despurinización espontánea del ADN.
 |
| Despurinización del ADN |
La luz ultravioleta podría provocar algo parecido. Esta
radiación provoca que se genere un dímero de pirimidina, se produce por
formaciónn de enlaces covalentes entre dos pirimidinas contiguas:
 |
| Dímero de pirimidian |
Al unirse, se crea una deformación del dúplex de ADN.
Quedarían mal emparejadas una serie de bases, opuestas al dímero. Si esto
sucediese durante la replicación, podrían cometerse errores. En concreto, se
cometería el mismo error que si faltase una base:
 |
| Dímero de pirimidina en el ADN |
Si se sustituyen un par de bases, esperamos que se vea
modificado, a los sumo, un aminoácido de la cadena. En la proteína, un solo
aminoácido se puede modificar por un error en una de las tres pares de bases
(recordemos que un aminoácido es codificado por tres pares de bases).
Por ejemplo, la secuencia AGC codifica para la serina. UGC
codifica para la cisteína. AAC codifica para la treonina. Y AGA codifica para
la arginina. Es decir, un cambio en una base puede provocar una sustitución por
uno de esos tres aminoácidos.
Corrimiento de la pauta de lectura.
Puede derivar de la inserción o deleción de un par de bases.
Un ejemplo es el que se produce cuando se inserta algún tipo
de compuesto policíclico que ocupa el lugar de una base, pero se empareja con
dos pares de bases.
En el hueco que queda debido a la inserción del compuesto
tricíclico podría insertarse una base más. Tras la replicación, en una de las
hebras habría un par de bases nuevas (en el dibujo, en la copia obtenida de la
hebra inferior).
La deleción es la pérdida de un par de bases. Por ejemplo,
cuando en el caso anterior, o en la formación de un dímero, en lugar de
completarse con una base de más, se suelta y se deja el hueco, quedando por lo
tanto una base de menos.
Debemos tener en cuenta que los corrimientos en la pauta de
lectura puede tener una trascendencia muy importante.
Veamos un ejemplo. la siguiente secuencia:
UAAUGCCGAAUCCGUACUAC
Codifica para la secuencia de aminoácidos:
Met-Pro-Tyr-Pro
Si se produce una deleción, en una de las bases de forma
que:
UAAUGCCGACAUUCCGUAC
Hemos pasado a la secuencia de aminoácidos:
Met-Pro-Thr-Ser-Val
Es decir, ha cambiado todos los aminoácidos que quedan por
delante del corrimiento de la pauta de lectura. A partir de la deleción (o de
la inserción) se cometen errores en todos los aminoácidos. Y la proteína no
será viable. Solo si quitamos o ponemos un número múltiplo de 3 pares de bases
la mutación puede no ser grave, a no ser que la mutación se compense más
adelante (en un lado quitamos una base y en otro lugar, más adelante, ponemos
una base de más, quedando el error o corrimiento acotado entre las bases del
medio).
Reparación de errores.
El simple hecho de que el ADN sea dúplex favorece una
facilidad para la reparación. Si se produce un error que afecte a una sola
hebra, la otra hebra siempre estará para poder reparar el error. El ARN no
tiene esta ventaja.
Un mecanismo de reparación es el que hay dentro del propio
proceso de replicación. En células superiores no se conoce el proceso exo-3’→5’. Pero teniendo en cuenta que el nivel de errores es
muy parecido al de las bacterias, se supone que debe existir.
El resto de errores se reparan por enzimas específicos. Los
enzimas reconocen el error, cortan la hebra que está mal, eliminan lo que está
mal y lo sustituyen. Después, cierran los extremos (con una ligasa).
Veamos el ejemplo con el caso del dímero de pirimidina. La
luz ultravioleta genera un dímero de pirimidina. Existe una endonucleasa que
identifica específicamente la deformación debida a este error, corta por un
lugar próximo al dímero, por el extremo 5’, a una distancia de unos seis
nucleótidos del error. Mediante la ADN-pol I se retira el trozo que tiene el
error y lo sustituye por el trozo correcto. Actúa como polimerasa 5’→3’ y como exonucleasa 5’→3’,
es decir, va limpiando el camino (debemos recordar que en la replicación quita
el cebador y ahora quita el error, es decir, es el mismo proceso pero
conceptualmente distinto).
 |
| Corrección del dímero de pirimidina |
Cuando hay bases equivocadas es necesario que actúe otro enzima.
Vamos a ver un ejemplo, cuando se sustituye citosina por uracilo. Necesitamos
un enzima que reconozca la base y la hidrolice, actuando después los tres
enzimas anteriores (endonucleasa, polimerasa y ligasa). El enzima que reconoce
el error por sustitución por uracilo es la Uracilo-DNA Glicidadasa.
 |
| Corrección de bases equivocadas |
Este sistema de reparación nos puede dar una razón de por
qué el sistema evolutivo ha seleccionado la timina en vez del uracilo y de por
qué no se siguió con el ARN como ácido nucléico (se cree que fue el primer tipo
de ácido nucléico). Si hubiese uracilo en vez de timina, no se podrían
distinguir los uracilos verdaderos de los falsos, es decir, de los derivados de
errores. Cambios químicos en el resto de bases originan bases extrañas que no
pertenecen al ADN. La Uracilo-ADN glicosidasa es específica para encontrar al
uracilo en el ADN y nunca lo confundirá con la timina.
Otra cuestión en los mecanismos de corrección es que los
enzimas de corrección sean capaces de distinguir la hebra progenitora, que actuó
como molde, de la hebra recién sintetizada. Es útil porque, si algo va mal, es
más fácil decidir cual es el error: la hebra molde será la correcta. La hebra
molde es reconocida porque está marcada, las adeninas están metiladas. Y tras
la replicación, el ADN tarda un tiempo en metilarse.
Consecuencias de las mutaciones.
Los errores que no se han corregido y que no se van a
corregir quedarán plasmados y ya no habrá posibilidades de corrección. Se ha
formado ya la mutación. Esta es la forma en que se producen las mutaciones.
Pero estas pueden tener diferente trascendencia. Dependerá de dos cosas: del
tipo de célula afectada y del tipo de mutación.
En cuanto al tipo de célula, es importante ya que la
mutación se trasmite a la descendencia, pero no a la descendencia del
individuo, sino a la descendencia de la célula. Podemos hablar de mutaciones
somáticas y de mutaciones germinales. Las somáticas se producen en cualquier
célula que no está implicada en la reproducción. La importancia de esta
mutación será a nivel de individuo, pero no se transmitirá a la descendencia.
Es una de las consecuencias del envejecimiento, la acumulación de mutaciones
somáticas. En cuanto a las mutaciones germinales, van a transmitirse a la
descendencia. Influyen en el acervo génico de la población. Son las más
importantes.
Según el tipo concreto de cambio, también podemos hablar de
diferentes tipos de modificaciones en el genotipo. Vamos a hablar de las
mutaciones según su trascendencia, de menos trascendentes a más trascendentes.
Por un lado hay mutaciones silenciosas, que no modifican en
absoluto el fenotipo. Es decir, el cambio no provoca modificaciones en
proteínas, aunque se modifica la secuencia de núcleotidos. Por ejemplo:
 |
| Mutación silenciosa |
En este caso la mutación no se manifestaría, ya que ambas
secuencias codifican para el mismo aminoácido.
Por otro lado tenemos mutaciones neutras. La modificación
del ADN sí va a variar el fenotipo, ero no va a variar la función de la
proteína. Es decir, la modificación del ADN provoca que haya un cambio en el
aminoácido de una proteína, pero este cambio no provoca cambio de función.
En el ejemplo, se cambia un aminoácido hidrófobo por otro y
poseen un tamaño parecido. Puede que en este caso no haya cambio en la
proteína, ya que es posible que no cambie ni la forma ni la función.
 |
| Mutación neutra |
Las mutaciones no tienen porque ser ni a favor ni en contra.
Y en estos casos de mutaciones neutras, no se apreciarían cambios. De hecho, en
una proteína no solo podrían ser inocuos cambios en un aminoácido. Podrían
incluso cambiarse dos o tres aminoácidos sin que sucediese nada.
Hablemos ahora de mutaciones visibles, que es lo que se
entiende por mutaciones propiamente dichas. Las mutaciones serán perjudiciales
en la mayoría de los casos. Esto no tiene una relación directa en que la
mutación se más amplia o menos. No tiene nada que ver con la cantidad de aminoácidos
que se modifican. Si se modifica un solo aminoácido, pero este pertenece al
sitio catalítico, podría desactivarse la proteína. Hay más probabilidad de que
esto ocurra cuantos más aminoácidos se modifiquen, cuantos más aminoácidos sean
modificados, más probabilidades hay de que afecten a la proteína, de que sean
perjudiciales y se obtenga una proteína que no es funcional. También puede
ocurrir que en el ARN mensajero, debido al cambiose bases, se obtenga una señal
de parada, con lo que se formarán polipéptidos en lugar de proteínas. Las
mutaciones perjudiciales van siendo eliminadas (selección natural).
También están las mutaciones beneficiosas, por ejemplo si se
forma un proteína más efectiva o que crea una función nueva, por ejemplo un
enzima con una especificidad distinta. En el primer caso, lo que suele suceder
es que el antiguo tipo de proteína va siendo sustituida por la nueva proteína
mejorada, la antigua va quedando obsoleta, la información va siendo sustituida,
hablándose entonces de evolución vertical. En el segundo caso solo puede
suceder que las dos proteínas coexistan, teniendo dos proteínas que nos
solucionan problemas y hablándose entonces de evolución horizontal. El ejemplo
más clásico de esto último es la hemoglobina, formada por dos tipos de
proteínas que si bien son distintas, provienen de una proteína ancestral común.
Pueden observarse familias de proteínas, comprobando como unas descienden de
otras.
 La luz solar contiene algunas radiaciones ionizasteis con cierta capacidad de provocar mutaciones. Las más energéticas, los rayos gamma y rayos X, se quedan atrapadas en la ionosfera. La radiación con mayor capacidad mutágena, sin embargo, es la radiación ultravioleta (UV), que llega en cierta proporción a la superficie terrestre.
La luz solar contiene algunas radiaciones ionizasteis con cierta capacidad de provocar mutaciones. Las más energéticas, los rayos gamma y rayos X, se quedan atrapadas en la ionosfera. La radiación con mayor capacidad mutágena, sin embargo, es la radiación ultravioleta (UV), que llega en cierta proporción a la superficie terrestre.