Aparato circulatorio.
Es un sistema abierto. Tiene un desarrollo inverso al
desarrollo de un sistema traqueal, está relacionado con el tipo de respiratorio
y circulatorio. Si la respiración es branquial o pulmonar, será complejo. Si es
traquial, el circulatorio no tiene función respiratoria, por lo que apenas
tendrá vasos y será sencillo.
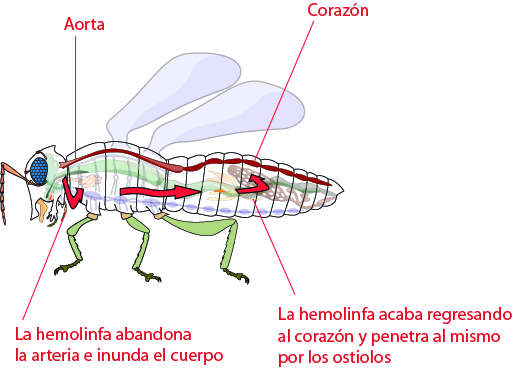
El corazón es un órgano musculoso, con musculatura estriada.
Y el tubo tendrá cámaras, entrículos, está metamerizado y está abierto a una
aorta posterior y a una aorta anterior. Entre cada ventrículo hay una válvula
para que no retroceda la sangre. Cada ventrículo tiene un par de ostiolos. Cada
ventrículo tiene un par de músculos laterales, los músculos alares.
 |
| Corazón en un artrópodo: sistema circulatorio abierto |
La
contracción de estos tira de las paredes del corazón, la distiende y la sangre
tiende a entrar tanto por la aorta posterior como por los ostiolos. El corazón
se contrae y la sangre sale disparada hacia delante. En caso de haberlo, la
sangre entra es del seno pericárdico.
 |
| Esquema del corazón del insecto |
La sangre va de atrás hacia delante. La sangre expulsada por
el corazón, por ejemplo, baña al cerebro.
La circulación tiene un sistema basado en corrientes de
líquido. Si el animal tiene un corazón y dos aortas hablaremos de corazón
aórtico, típico por ejemplo de los insectos. Su función no es respiratoria. En
el caso de los crustáceos, que tienen branquias, o pericelados con pulmones,
nos aparecerán vasos que recorren el cuerpo del animal. Los más típicos van
desde el respiratorio hasta el corazón. Pueden ser vasos, rodeados de epitelio,
o senos. Cuándo el animal es muy grande o con actividad muy potente, habrá
venas desde algunas partes del cuerpo al órgano respiratorio. A los sitemas de
los animales con respiración por pulmones o tráqueas se les llama sistemas
venosos.
 |
| Circulatorio en insectos |
La composición de la sangre es muy variada, tanto el plasma
como los tipos celulares. El único que es casi general son los granulocitos,
encargados de fagocitar invasores y coagular la sangre. Pueden aparecer o no
pigmentos respiratorios. En caso de aparecer, son funamenalmente dos, la
hemocianina, en crustaceos y arácnidos y la hemoglobina, en algunos crustaceos
y algunos ejemplos puntuales de insectos. Cuando aparece hemoglobina en
insectos es porque tienen una vida en zonas con gran escasez de oxígeno.
La principal función de la sangre en la mayor parte de los
artrópodos, no obstante, es el transporte de sustancias entre las partes del
animal.
Aparato excretor.
En el aparato excretor encontramos dos tipos de órganos, los
celomiductos y los tubos de Malpighio. Los celomiductos son la herencia de
grupos anteriores, con nefridios modificados. Los tubos de Malpighio son
órganos nuevos. Los artrópodos deben expulsar los productos derivados del
metabolismo del nitrógeno o retener o wliminar agua o iones, dependiendo de los
niveles internos en cada caso.
Celomiductos.
Su nombre hace referencia a su origen, es decir, conductos
derivados del celoma. Tiene dos partes, un sáculo y un conducto denominado
celomiconducto. El celomiconducto se abre al exterior por un poro. En la zona
final del tubo puede haber una vejiga.
En el sáculo, que conserva uno de los pocos restos del
celoma, tiene lugar la ultrafiltración. En el celomiducto se resorben los
iones, agua y aminoácidos. El poro excretor se abre al exterior y se abre a
diferentes sitios del cuerpo dependiendo del grupo. En crustaceos, en la base
de las segundas antenas o en las segundas axilas. También se abre ahí en sus
larvas, la larva nauptilus, pero de forma contrario al adulto, es decir, si en
el adulto está en las axilas, en la larva estará en las antenas. En quelicerados
se encuentra en las coxas. En anterocerados, en la base de algún apéndice
bucal. En estos últimos están poco desarrollados, estando mucho más
desarrollados los tubos de Malpighio.
Tubos de Malpighio.
Siempre están relacionados con el tubo digestivo, sus
productos de excreción van a parar a éste. Son unos tubos compuestos por un
epitelio con microvellosidades. A veces tiene alguna fibra muscular (siempre
pocas, en el caso de que las haya). Estas fibras musculares permitirían la
movilidad del tubo, que ondea por el interior de la hemolinfa.
 |
| Tubos de Malpighio |
El tubo tiene una zona distal y una zona proximal. En la
distan tiene lugar la ultrafiltración, se filtra la hemolinfa con sus
componentes. El epitelio de esa zona está repleto de microvellosidades. En la
zona proximal tiene pocas microvellosidades y el sentido de la filtración es el
contrario, reteniendo agua, iones, etc. El tubo se va cargando progresivamente
de productos del metabolismo del nitrógeno.
El desarrollo y el número de tubos varía en los diferentes grupos.
En crustáceos solo van a aparecer, en algunos, rudimentos de este tipo de
tubos, en forma de papilas en el recto. El quelicerados terrestres su
desarrollo es más potente y se abren al final del mesodeo, antes de la válvula
pilórica.
En los aracnidos van a tener mucha importancia, están
desarrollados y se abren al principio del proctodeo. Las pequeñas cantidades de
agua que se pierden en la orina de estos animales es recuperada en el recto,
llegando a resorberse casi el cien por cien del agua.
Nos vamos a encontrar con distintos tipos de excreción.
Algunos son aminotólicos, como los crustaceos. Otros producen ácido úrico, como
los mirápodos y los insectos. En la zona intermedia estará la urea, que es poco
tóxica y bastante soluble. Aparece en animales que viven en lugares con mucho
vapor de agua y nunca aparecerá como sistema de excreción única. Lo presentan
algunas arañas, que también segregan urea y guanina, por ejemplo.
Nefrocitos.
En los crustáceos se colocan n los ejes de las bránqueas. A
veces funcionan como riñones de acumulación, son fagocitarios y después
expulsan lo fagocitdo. También pueden funcionar como centros metabólicos,
degradando parcialmente sustancias complejas que captan de la hemolinfa.
Están desarrollados en larvas y en pupas. Acumulan lo que fagocitan los
nefrocitos y después se desprenden con la cutícula en la muda imaginal.
Sistemas respiratorios.
No hay un modelo para todos ellos. Se cree que los
artrópodos primitivos tenían respiración cutánea. La cutícula húmeda es muy
permeable a los gases. Un animal de pequeño tamaño, si tiene la cutícula
humedecida, puede respirar por ella.
En algunos animales suponemos que toda la superficie, todo
el tegmento, respira. No hay estructuras especializadas. Por ejemplo, los
picnogónidos.
Si el animal aumenta de tamaño o tiene una actividad
importante, la respiración cutánea no erá suficiente, se necesita más oxígeno y
eliminar más dióxido de carbono. Cuando eso ocurre, hay áreas de respiración
cutánea especializadas. Estas van a cumplir por lo menos una condición: el
tegumento debe estar adelgazado con respecto al resto del cuerpo. El
intercambio va a aumentar en esa zona.
Además suelen ocurrir otros dos fenómenos. Suelen aparecer
apéndices ventiladores del área, que renuevan el medio externo, sea éste aire o
agua, de forma que eliminan las concentraciones altas de dióxido de carbono que
se acumularían y pasaría medio fresco, rico en oxígeno. E internamente, la
hemolinfa suele ponerse en movimiento, apareciendo un sistema de circulación.
De este sistema de respiración derivarán todos los demás. En
esta línea se ha evolucionado de distintas maneras. Van a aparecernos branquias
o pulmones. En ambas ocurre lo mismo, es decir, un aumento de la superficie
cuticular por replegamiento de esta. Mantienen una pequeña porción de
superficie adelgazada y la aumentan por evaginación o invaginación, dando lugar
a branquias y pulmones respectivamente.
El otro aparato de respiración es el traqueal. Es una región
que se invagina. Comienza formando estructuras pulmonares o asimilables a
pulmones en las menos evolucionadas. Estas invaginaciones se hacen muy
desarrolladas, toman forma de árbol y la punta más fina de la rama está en
contacto con un pequeño grupo de células, con las que realiza el intercambio.
Es decir, el intercambio no tiene nunca lugar con la hemolinfa. Siempre hay un
pequeño intercambio entre el aparato traqueal y la linfa, pero es mínimo, solo
sirve para que respiren las células de la hemolinfa y poco más.
Respiración cutánea.
La presentan los picnogónidos, crustáceos pequeños y larves,
alguna apteriogotas, ácaros, algunos paurápodos, etc. La respiración cutánea
más especializada aparece en los percebes (cirrípedos).
Branquias.
En todos los crustáceos se suelen asocia las branquias con
unos apéndices, pues los apéndices están en movimiento y así se airea. Estas
branquias presentan una complejidad variable, desde hojas sobre los apéndices
hasta estructuras mucho más complejas.
Los decápodos las tienen sujetas a una pata y con una
circulación por senos sanguíneos. Está protegido por un caparazón y tiene una
cámara branquial. El animal tiene un agitador en el esoafagmato, que hace que
circule el agua. En el género Macrura se complica un poco (son las langostas,
cangrejos de río y son diferentes a las Metantia, que son las gambas),
cerrándose el caparazón y estando solo abierto por la base de las patas, para
poder caminar. En esa zona se encuentra la entrada de agua. El caso más
complicado es el del género Brachyura, que son los cangrejos auténticos. En
ellos todo el caparazón se funde con la parte central del cuerpo. Centralmente
ya no hay orificios. Desarrollan un par de poros en posición anterior ventral.
El agua entra por uno de esos poros, da vuelta y sale por el otro. Cuando las
branquias se van tapando por impurezas, invierte el sentido y así arrolla todos
los depósitos. En algunos hay una especie de látigo que se encarga de limpiar
las branquias.
Hay más artrópodos que presentan branquias, pero no los
veremos.
Existen animales con respiración branquial que se han
adaptado a la vida terrestre, como el cangrejo de los cocoteros (un cangrejo
especializado en trepar por los cocoteros y romper el peciolo del coco). Este
vive en tierra casi todo el tiempo. Las laminillas branquiales están mojadas y
respira tranquilamente. Su principal problema es la excreción, las branquias
eliminan amoniaco y debe meterse en el agua para lavarlas y evitar envenenarse.
Posteriormente vuelve a salir, siguiendo con su vida terrestre.
Pulmones.
Han evolucionado de forma independiente en miriápodos,
quelicerados terrestres y crustáceos.
 |
| Pulmón en libro (aracnidos). |
En Scutiqueracela optrata en cada tergo hay un poro
anterior, exhalante, y un poro posterior inhalante. Por movimientos del tergo
logramos que el aire entre por un lado y salga por otro. El aire entra en el
atrio colocado debajo del tergo, sobre el seno pericárdico. Al seno pericárdico
envía unos tubos, que ya no son branquias. El intercambio tiene lugar entre
esos tubos y la hemolinfa del seno pericárdico. Esa hemolinfa pasa al corazón
por los ostiolos y sigue su circulación. Es un sistema exclusivo de
Scutriromorfos. Es el único miriápodo con aparato pulmonar. El resto posee
sistemas prototraqueales o traqueales, con tubos que penetran en la hemolinfa.
En los Sifonuros, los apéndices pequeños de la parte de
atrás están replegados, constituyen branquias, con forma de hoja, apretadas,
unidas. De ellas derivan los escorpiones. En estos, se han convertido en
pulmones. Nos aparecen en el embrión del animal en forma de branquias, pero en
el interior. Se forman una serie de hojas bañadas por hemolinfa. Este tipo de
pulmón lo tienen los policerados. Las láminas de las branquias tienen puentes
que las separan, para que no se peguen unas a otras. A vences aparecen músculos
encargados de ventilar la zona.
En los escorpiones aparecen cuatro pares de pulmones. En los
arácnidos, dos pares en los más primitivos, que son reemplazados por tráqueas
en los más evolucionados. En estos presentan dos pares de tráqueas, que son más
eficaces.
Hay otro sistema pulmonar, el de los crustáceos terrestres e
isópodos terrestres. Algunos apéndices son aplanados y aparece en su cara
interna un poro, denominado expirópodo, que comunica con una red de tubos
huecos que se llenan de aire y que forman el sistema pulmonar denominado
pseudotráquea. No es una tráquea, ya que intercambia gases con la hemolinfa, no
con los tejidos. Los isópodos tienen un sistema branquial, parte del apéndice
funciona como branquia, tiene estrías con microestrucuturas que son capaces de
condensar la hemolinfa y bañan esa parte branquial. De este modo está siempre
mojada y funciona como una branquia. Ese mismo sistema sirve para que el animal
desarrolle sus embriones, pues tiene una cámara llena de agua.
En los animales de sistema respiratorio pulmonar el sistema
circulatorio debe estar muy desarrollado.
Sistema Traqueal.
Presente en artrópodos terrestres, incluso en arañas.
Consiste en una red de tubos huecos que llevan el aire a todos los tejidos. En
la hemolinfa no habrá pigmento respiratorio. El sistema traqueal no suele tener
dos órganos diferentes, las tráqueas y las traqueolas. Las tráqueas disminuyen
de diámetro y tras ellas se colocan las traqueolas, pero como estructuras
difernciadas.
El más desarrollado aparece en insectos. Las tráqueas tienen
una pared igual al tegumento externo, pero invertido, es decir, la cutícula
está por dentro. Toda la tráquea tiene un engrosamiento en espiral, el
ctenidio, que evita que la tráquea se colapse con facilidad, siendo físicamente
más efectivo. La exocutícula le da rigidez a la tráquea.
 |
| Sistema traqueal eninsectos |
En algunas zonas se pierde la exocutícula, y en esa zona el
tubo es más flexible. Hablamos entonces de sacos aéreos. Sirven para que el
animal aumente de volumen tras la ecdisis. Otra función es ventilar el sistema
traqueal y emitir o recibir sonidos. Además, pueden contribuir al vuelo, cuando
se va a volar se coloca perpendicular al sol naciente para calentar el aire del
saco aéreo, consiguiendo que el aire caliente ayuda a subir.
Las tráqueas se comunican con el exterior por unos orificios
llamados estigmas o espiráculos. Los troncos traqueales que parten de cada
esperiáculo, y que suelen corresponder a dos por metámero, en artrópodos
inferiores no se conectan unos con otros. Según la evolución avanza se unen
para que el sistema trabaje junto, entrando todo el sistema en contacto. Se
conectarán tanto dorsal como centralmente, por todas partes.
Este sistema, en las ramificaciones más finas, son las que
se conocen como traqueolas. En ellas encontramos los traqueoblastos, células
muy ramificadas y que llevan una red de canales intracelulares. Estos tubos
internos no tienen epicutícula, solo una fina capa de procutícula. No hay
tenidios y aparecen unos espaciamientos que le dan un poco de rigidez. Las ramificaciones
del traqueoblasto se colocan sobre el tejido que van a airear y como la pared
es muy fina, es permeable a los gases.
Cuando el tejido está en reposo, las traqueolas se llenan de
líquido intracelular. Cuando el tejido entra en actividad, el líquido se
reabsorbe y las traqueolas se llenan de aire, produciéndose el intercambio de
gases. Dependiendo del tejido, puede haber más o menos traqueolas con más o
menos traqueblastos.
Los sacos aéreos intervienen en la ventilación. Cuando el
artrópodo está en actividad, el animal, por movimientos del abdomen, aplasta o
estira los sacos aéreos, puede aplastar o expandir el abdomen, hinchando o
aplastando con ello los sacos aéreos.
Los insectos acuáticos tienen sistema traqueal. El más
sencillo es tomar aire acudiendo a la superficie, pero tiene el problema de que
los expone a los depredadores. Algunos insectos han desarrollado tubos
respiratorios que los ponen en contacto con el exterior, con la superficie. En
otros casos, al animal toma una burbuja de aire bajo las alas, enganchadas a
pelos, y respira de esa burbuja, pero aunque entra algo de oxígeno del agua, se
le va mucho, la burbuja disminuye y tiene que salir a la superficie a
renovarla. El problema lo han resuelto los insectos que son capaces de tomar oxígeno
del agua a través de las tráqueas. Hay dos sistemas, las traqueobranquias y las
plastrón.
Las traqueobranquias aparecen siempre en larvas. Los
animales tienen cerrados todos los estigmas casi siempre. Están conectados con
las traqueobranquias. El estigma se cierra por una estructura que la separa de
traqueas ramificadas del exterior. Estas estructuras de cierre tienen una
cutícula muy fina y húmeda, de tal forma que el aire de la traquea intercambia
gases con el agua del exterior. Se trata de estructuras muy delgadas y
sobresalientes, que tiende a romperse. En anisópteros los han desarrollado
dentro del tubo digestivo y el animal respira intermitentemente.
 |
| Traqueobranquia. |
El sistema más sofisticado son los plastrones. Son una capa
de pelos hidrófobos muy cortos, de unas pocas micras, y muy apretados, llegando
al millón por milímetro cuadrado. Habrá espiráculos abiertos. El aire de la
traque llega al exterior. Los pelos guardan una fina capa de aire, una lámina
de aire. En este caso no ocurre como en la burbuja, no se pierde aire porque
los pelos lo agarran y la capa de aire intercambia oxígeno y dióxido de carbono
con el agua continuamente. Impide el intercambio, pero no con la salida del
volumen de agua. Cuando el animal bucea, la burbuja se aprieta más al cuerpo porque
se pegan los pelos. En realidad siempre se pierde algo de gas, pero puede durar
varios días. En los más complejos, es eterno, no se llega a gastar, pues los
ápices son paralelos a la superficie del cuerpo.